Article Text

Abstract
SARS-CoV-2 infection can have widely diverse clinical outcomes, from asymptomatic infection to death, with many possible clinical symptoms and syndromes. It is thus essential to understand how the virus interacts with the host immune system to bring about these varied outcomes and to inform vaccine development. We now know that both antibody and T cell responses are induced in the majority of infected individuals, and that cross-reactive responses from other coronaviruses also exist in the uninfected population. Innate immune responses are a key focus of research and may influence the course of disease and the character of subsequent adaptive responses. Finally, baseline immune profiles and changes during early acute infection may be key to predicting the course of disease. Understanding all these aspects can help to create better immune monitoring tools for COVID-19, including tools for predicting disease severity or specific sequelae, perhaps even prior to infection.
- adaptive immunity
- inflammation mediators
- immunity
- cellular
- immunity
- humoral
- immunity
- innate
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
SARS-CoV-2 can cause asymptomatic infection in some individuals, mild-to-moderate self-resolving symptoms in others and severe manifestations like acute respiratory distress syndrome (ARDS), coagulation disorders and death in smaller subsets of patients.1–3 The range of symptoms is highly variable and can include many different organs and systems.4 In addition, some individuals experience prolonged symptoms, if not prolonged viremia, a phenomenon often referred to as ‘long COVID-19’ or ‘postacute COVID-19’.5 Among children, a small proportion develop a multiorgan inflammatory syndrome (MIS-C),6 7 usually after resolution of viral infection. While clinical factors such as age and pre-existing conditions like obesity and diabetes are known to increase the risk of severe disease,8 9 there is an inability to accurately predict who is at greatest risk, and who will experience particular symptoms or sequelae.
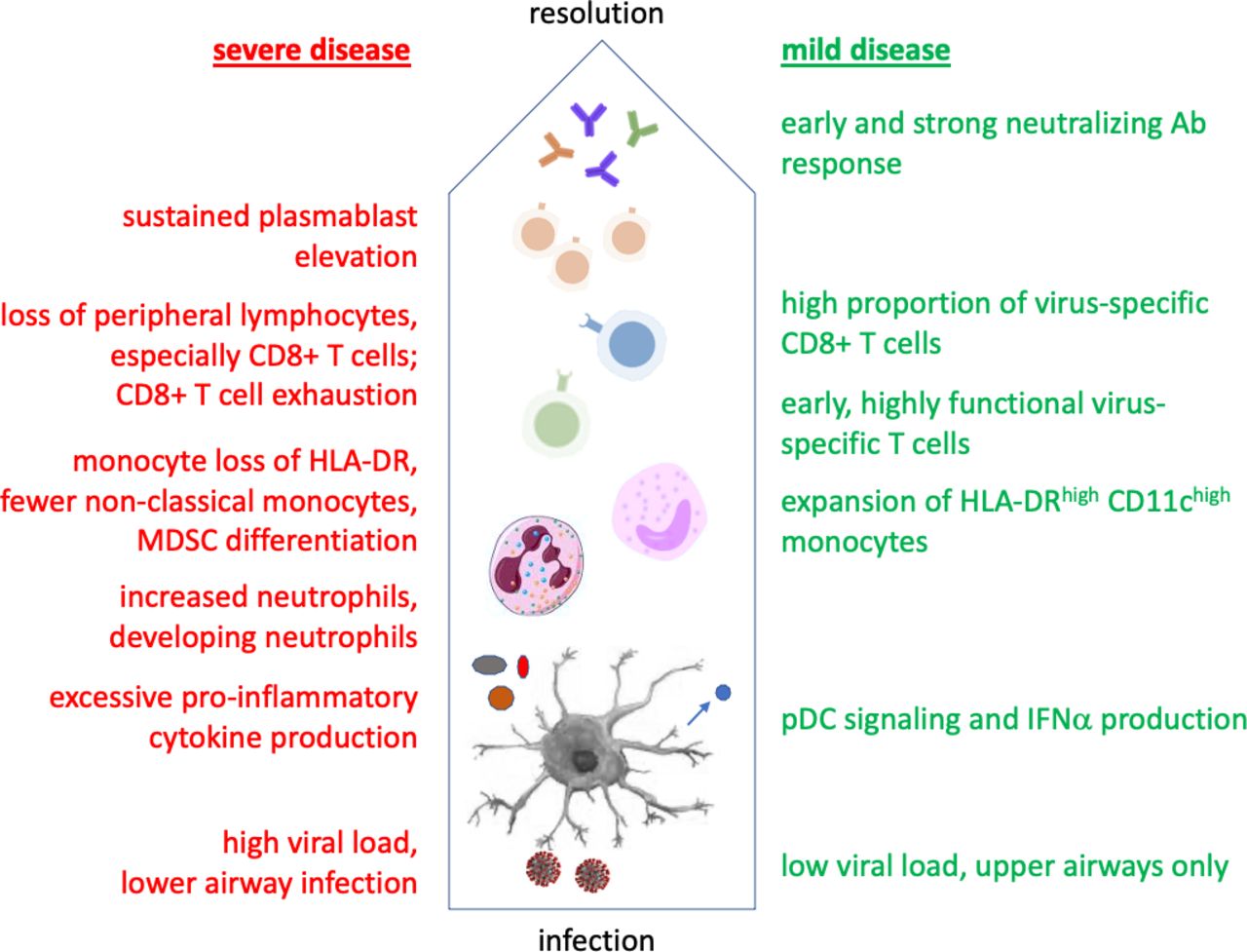
Variation in the spectrum of COVID-19 disease could potentially derive from viral variation, which is increasingly being described.10 11 However, it is not yet clear if emerging mutant virus strains cause increased disease severity, or simply increased transmissibility. There can certainly be a contribution of host genetic factors, including some related to viral binding and entry.12–14 But genetic variation in the immune response has also been implicated, including recently discovered variants in type I IFN immunity associated with severe disease.15 In fact, variability in the host immune response is likely to play a major role in determining the course of SARS-CoV-2 infection, as exemplified in figure 1 and discussed in detail here. Given this, there is a need to apply immune profiling to better understand the interaction of this virus with the human immune system and how this influences clinical outcomes and informs vaccine development.

Immunological course of mild versus severe COVID-19. Cells and secreted proteins involved in the immune response are depicted from the bottom up, beginning with viral interactions with antigen-presenting cells such as DCs. Findings in green versus red have been associated with mild versus severe disease, respectively. DCs, dendritic cells; MDSC, myeloid-derived suppressor cells; pDC, plasmacytoid.
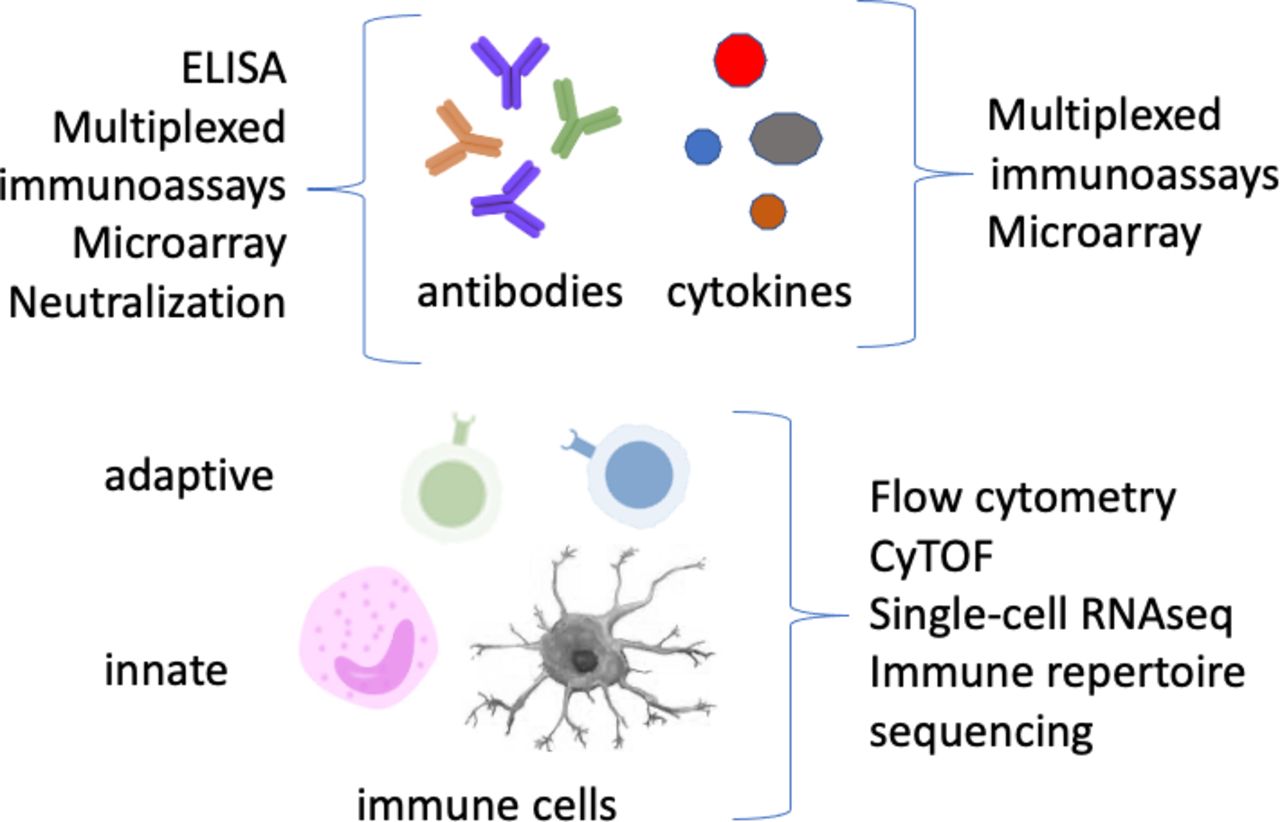
While other papers in this review series will address the interaction of cancer and COVID-19, this review will provide an overview of COVID-19 immune profiling in general. Immune profiling is usually divided into humoral or serology assays, which measure antibody responses and soluble factors such as cytokines; and cellular assays, which assess global as well as antigen-specific immune cells (figure 2). These latter assays can profile cells of the innate and/or adaptive immune system. All of these immune responses are relevant to COVID-19, so while they are reviewed individually here, a comprehensive approach will be most informative. For each type of immune finding, one or a few key references are provided, since the literature is already so vast as to preclude citing all relevant papers in this field. In addition, this review is focused mostly on immune responses as measured in blood; but valuable observations (discriminating COVID-19 severity levels, eg) can be made from single-cell studies of tissues such as lung, or fluids such as bronchoalveolar lavage, when available.16 17
{kind=link}
{kind=link}
Immunological biomarkers and common assay platforms employed to detect them. Techniques listed are not meant to be exhaustive.
Serology: antibodies
Antibody responses are the most-studied aspect of the immune system, and have certainly been well characterized in COVID-19.18 They can be measured by ELISA as well as by multiplexed immunoassays, using bead-based fluorescent or microtiter plate electrochemiluminescence readouts. Even higher multiplexing can be achieved with protein/peptide microarrays.19 20 Neutralization capacity can be measured by infectivity assays, for which lentiviral pseudoparticles expressing SARS-CoV-2 spike protein have been employed to reduce biohazard risks.21
Most infected individuals raise a detectable antibody response to SARS-CoV-2, including IgM, IgG, and IgA antibodies to the spike protein and its receptor-binding domain.18 22 These responses tend to be higher in more severe cases, though there is considerable population heterogeneity. Antibody levels generally peak within 2 weeks for IgM and 3 weeks for IgG,23 for both nucleocapsid and spike protein targets. These responses are relatively stable for at least 8 months or more, though the degree of decline has been debated. While one large Islandic study24 and another even larger New York study25 reported antibodies ‘do not decline’ and ‘persist’ over several months, other (smaller) studies showed a slow decay in titers,26 27 with one study showing a median decrease of 34.8% over 3 months.26 These observations were also more or less corroborated by a preprint describing a very large UK cohort, where a mean decrease of 26.5% over 3 months was observed.28 This decline was age-dependent, with larger decreases in the elderly. Another healthcare worker study reported that 91% of seropositive individuals remained seropositive over 3–5 months, which is in keeping with generally small declines over this period.29 A good correlation between total and neutralizing IgG antibodies has also been reported.26
Interestingly, memory B cells seem to persist or increase even as antibody levels wane.30 31 Also, Gaebler et al32 showed that memory B cells not only persist, but undergo somatic hypermutation over the course of 6 months, consistent with persistent antigen.
What is less certain is whether protection against COVID-19 is durable in a time frame of years, given that protection against other circulating coronaviruses tends to be short lived, as seen by increased reinfection after 1 year.33 This could be due to waning antibody titers, particularly neutralizing antibodies, and/or declining T cell responses, the latter to be discussed further on in this review. Interestingly, germinal centers are significantly blocked in COVID-19, which might affect antibody quality and durability.34 Nevertheless, somatic hypermutation of antiviral antibodies still occurs.30 32 Cagigi et al35 also found that airway antibodies tend to wane quickly, despite persistent serum IgG in patients with COVID-19.
All these studies prompt questions about the durability of vaccine responses, as COVID-19 vaccines are now being distributed at an unprecedented rate. Fortunately, the data to date show no drastic decline in antibody titers to day 119 in recipients of an mRNA vaccine.36 Whether an annual ‘booster’ will be needed is still debated, but it’s likely that circulation of new variants will drive this need at least as much if not more than waning antibody titers.
Despite the short-lived protection against seasonal coronaviruses, many people (especially children) harbor antibodies to one or more of these, and such antibodies can be cross-reactive with SARS-CoV-2.19 22 37 This could account for some of the decreased COVID-19 severity generally seen in children. However, children (like adults with mild or asymptomatic disease) tend to produce lower antibody responses to SARS-CoV-2 once infected.38
Mapping the protein or even epitope specificity of antibody responses could provide potential correlates of protection or disease severity. One report showed nucleocapsid-specific IgG to be significantly higher in intensive-care unit (ICU) versus non-ICU patients, while spike-specific IgG was higher in non-ICU patients.23 The combination of nucleocapsid-specific and spike-specific antibody detection was also helpful in identifying more infected patients (75% with the combination).
Some studies have speculated on the value of serology against heterologous antigens in the context of COVID-19. Vaccines such as diphtheria-tetanus-pertussis share some cross-reactive B cell epitopes with SARS-CoV2,39 which could offer a level of cross-protection in children; but this idea is so far untested. CMV infection could contribute to immunosenescence and the general severity of COVID-19 in the elderly,40 though this too is speculative. For influenza, there is little epitope cross-reactivity with SARS-CoV-2,39 though there is one report showing a reduced incidence of COVID-19 respiratory symptoms in pediatric patients vaccinated for influenza versus those not vaccinated.41 Of course, this could also be due to confounders not controlled for in the study.
From these observations, we can derive recommendations for prospective studies to employ a comprehensive serology assessment. This should include multiple proteins/epitopes of SARS-CoV-2, to maximize identification of seropositives and potentially detect differences between severity groups. Inclusion of heterologous antigens, such as those from seasonal coronaviruses, could also be informative.
Serology: cytokines and other soluble mediators
Cytokines are important signaling molecules of the immune response, and their elevation in blood or in nasal swabs or aspirates may be predictive and/or correlative with the course of disease. Platforms for measuring soluble cytokines have evolved from traditional ELISA to multiplexed platforms such as fluorescent bead-based assays, electrochemiluminescence assays, or microarrays. Proximity extension assay (PEA) can also be used for multiplexed readout of cytokines and other soluble proteins. PEA employs pairs of DNA-tagged antibodies to generate a polymerase chain reaction (PCR) product that is read out by quantitative PCR (qPCR). This technology provides very high multiplexing with good specificity,42 and is being used in large COVID-19 clinical studies such as IMPACC (https://www.niaid.nih.gov/clinical-trials/immunophenotyping-assessment-covid-19-cohort-impacc). It has also been used extensively in systems-level COVID-19 studies such as Su et al.43 An even greater level of multiplexing has been achieved with aptamer-based assays, which have also been used to identify host proteins, many of them associated with a genetic variation that interact with or are involved in a maladaptive response to SARS-CoV-2.44
A key feature of severe COVID-19-associated ARDS is the high level of many proinflammatory cytokines in serum, a so-called ‘cytokine storm’.45 While controlling these cytokines has been thought by many to be important,46 there is controversy about the relevance of these cytokines to disease pathology, and whether the ‘storm’ is even as severe as in other syndromes like sepsis-related ARDS.47 48 Nevertheless, it is clear that serum cytokine levels are a predictor of disease severity, including interleukin (IL)-6, IL-8, and tumor necrosis factor (TNF)49 and growth/differentiation factor-15 (GDF-15).50 Serum levels of TNSF14 (LIGHT), EN-RAGE (an inflammatory cytokine associated with coronary heart disease), and oncostatin-M (an IL-6 family member) have also been correlated with disease severity.51 Clinically, studies have targeted granulocyte/macrophage-colony stimulating factor (GM-CSF)52 53 and IL-154 with monoclonal antibodies and seen clinical benefit in patients with severe COVID-19 and ARDS. Zhao et al have also shown that high levels of serum GM-CSF and IL-17A correlated with more severe disease, and were linked to lung-resident Th17 cells.55 While all of these can be considered proinflammatory cytokines, together they represent a similar but not necessarily identical set of cytokines compared with those elevated in sepsis and other respiratory illnesses.
Other soluble protein biomarkers have also been identified as elevated in COVID-19. These include long pentraxin 3, expressed by monocytes and endothelial cells, and whose plasma level was found to be predictive of mortality in hospitalized patients with COVID-19.56 Similarly, plasma levels of calprotectin, together with non-classical monocyte frequencies, could be predictive of those who develop severe COVID-19.57 58 In fact, a signature of multiple soluble biomarkers is probably ideal for predicting disease course. Abers et al59 described a set of 12 soluble proteins, including various cytokines, whose initial level on hospital admission was predictive of mortality. Four of these (sTNFRSF1A, sST2, IL-15, and IL-10) longitudinally separated survivors throughout the course of hospitalization.
Given the somewhat eclectic list of cytokines associated with COVID-19 severity, there is as of yet no single panel that uniquely covers all of these. Rather, most groups continue to use highly multiplexed research platforms. Alternatively, a simple real-time readout of a few key predictive cytokines could be used for early patient assessment, as was done by Del Valle et al49 using a commercial point-of-care immunoassay system.
Cellular immune monitoring
Cellular immunity is inherently more complex to analyze, but new technologies have broadened our ability to do complex immunophenotyping at the single-cell level. Standardized immunophenotyping panels have been described for high-parameter flow cytometry60 and Cytometry by Time of Flight (CyTOF) mass cytometry.61 62 Many studies also use single-cell transcriptomics, which can be adapted to include staining with DNA-tagged antibodies (AbSeq or CITE-seq),63 as well as targeting of rearranged T cell receptor (TCR) and Ig genes. This obviously creates a powerful, unbiased approach for single cell phenotyping, but one which still suffers from issues of high cost and low throughput, as well as analytical complexity. Another single-cell technology, employing microwell-based cytokine detection to quantify the ‘secretome’ of up to 32 cytokines secreted by activated T cells, has also been used in multiomics studies to distinguish mild from moderate or severe COVID-19.43
Innate cellular immunity
Recent years have seen a greater appreciation for the key role of innate immune cells, which encompass granulocytes (neutrophils, eosinophils, and basophils), monocytes and macrophages; dendritic cells (myeloid and plasmacytoid (pDCs)); and innate lymphoid cells (ILCs, which include natural killer (NK) cells). While these cells do not have antigen-specific receptors, they can in some cases develop a heightened responsiveness, or so-called ‘trained immunity’ to certain pathogens.64 And, of course, the innate response triggers, and its character helps determine, the magnitude and quality of the ensuing adaptive response.
Trained immunity, including that induced by the tuberculosis vaccine BCG65 may influence the susceptibility of different populations to COVID-19 and/or to severe disease.66 Such differences in innate immunity may underlie geographical differences in overall severity of COVID-19 and could be part of the protection from severe disease generally seen in children.
Among patients with COVID-19, a major phenotypic finding of innate immune cells is the down-regulation of human leukocyte antigen-DR isotype (HLA-DR) on monocytes, which appears to correlate with severity.51 57 67 68 This could be related to early virus-induced type I IFN signaling, which has been a subject of study in COVID-19 and influenza.69 In severe COVID-19 cases, decreased IFNα production and reduced mTOR signaling in pDCs has been described.51 This is reinforced by the frequent finding of inborn errors of type I IFN signaling15 or autoantibodies to type I IFN70 in patients with severe COVID-19. Interestingly, the German DeCOI consortium found an increase in HLA-DRhigh CD11chigh monocytes with a strong IFN-stimulated gene signature in patients with mild COVID-19.67 In severe cases, these cells were absent, and the authors instead observed HLA-DRlow monocytes. A recent study also found myeloid-derived suppressor cells (MDSCs) in the circulation (but not airways) of patients with COVID-19, in a severity-dependent manner.71 In fact, many studies have reported a range of changes across the myeloid compartment in COVID-19.51 57 67 68 72 73
Neutrophils are also affected in severe COVID-19, as a result of emergency hematopoiesis generating circulating neutrophil precursors, or developing neutrophils,67 74 which have a unique gene expression profile that is similar to that of plasmablasts.74
NK cells are activated in COVID-19 as well, and distinct immunotypes of NK cells can be linked with disease severity.75 Additionally, mucosal-associated invariant T cells (MAIT cells), an innate-like T cell population, have been shown to both decrease in number and show an activated phenotype but impaired function in patients with COVID-19.76 77 Interestingly, the level of HLA-DR expression on MAIT cells was found to correlate with a disease severity score.76
Adaptive cellular immunity
Adaptive immune responses are those elaborated by B and T cells, which uniquely expand clones of memory cells with receptors (Ig and TCR) specific for pathogen epitopes. Bulk or single cell sequencing of those receptors allows the identification of expanded clones. The antigenic specificity of those clones remains difficult to infer, though computational methods for this have been developed.78 T cell specificity can also be screened by stimulation with peptides, often using overlapping peptide pools, which can be deconvoluted to determine individual peptide reactivity. The responses are generally read out by flow cytometry (with intracellular cytokine and/or activation marker staining), or by ELISpot assays, which measure single-cell cytokine secretion in a 96-well plate format. Interestingly, T cell responses in unexposed individuals were preferentially seen with overnight, rather than longer (6 d) stimulation,79 highlighting the importance of assay conditions in these types of experiments.
SARS-CoV-2 infection results in detectable B and T cell responses in the vast majority of cases, with generally robust responses in mild infection.37 80–88 T cell responses have even been described in close contacts of infected individuals, in the absence of detectable viral infection.89 It is not yet clear whether differences in protein targets and epitope specificity may be associated with severity; however, children tend to have lower magnitude responses to spike protein, despite their relative lack of severe disease.90 Grifoni et al84 showed that 70% of convalescent patients had CD4+ T cell responses, while 100% had CD8 +T cell responses to SARS-CoV-2,84 using predicted epitope peptide pools. These were further mapped to show a wide breadth of protein target specificities. Peng et al80 further showed that mild cases had higher proportions of virus-specific CD8 +T cells.
Recently, Tan et al91 showed that early induction of IFNγ-producing T cells responding to SARS-CoV-2 proteins was associated with viral clearance and milder disease. Thus, the kinetics of T cell responses could be important in disease course, and early measurement could help predict severity.
The clonal distribution of the T and B cell responses to SARS-CoV-2 may also play a role in determining disease course. Schultheiß et al92 have created a public database of over 14 000 000 B cell receptor and TCR sequences from the blood of acute and convalescent patients with COVID-19. TCR clonality and skewing were associated with flow cytometric findings of T cell activation phenotypes and interferon responses. Certain TCR clones were exclusively associated with recovery. Cheng et al93 have also studied TCR clonal expansions in adult patients with COVID-19 with hyperinflammation, and find TCR skewing that may result from superantigen-like interactions of the SARS-CoV-2 spike protein with MHC class II.
Multiple studies have also characterized T cell phenotypes and functions in the context of COVID-19.37 80 81 85 87 94 These tend to show a robust and highly functional T cell response, with a Th1 phenotype and good proliferative potential, following mild to moderate disease.85 87 While severe disease can be associated with a higher proportion of SARS-CoV-2-specific T cells,80 they may have a more exhausted phenotype,82 with less functionality and a lower IFNγ/TNFα ratio compared with influenza-specific T cells.
As with antibodies, T cell responses can be cross-reactive with other common coronaviruses. This can be seen in the form of SARS-CoV-2-reactive T cells in the blood of uninfected individuals.30 37 84 85 94 These cells tend to have lower avidity for SARS-CoV-2, but likely correspond to a predisposition toward milder disease should these individuals become exposed to SARS-CoV-2.
Additionally, Paschold et al95 found IgH gene rearrangements associated with SARS-CoV-2 reactivity in the repertoire of prepandemic individuals’ B cells. These were of particular interest, as they were decreased in individuals over 60 years old and in those with cancer—potentially accounting for some increased risk of severe COVID-19 in these groups.
Broad immunophenotyping
While studies of antigen-specific T and B cells can be highly informative, these are rare populations and assays to identify them are more complex than simple immunophenotyping assays. But important information can also lie in the proportions and phenotypes of broader cell populations. This includes total lymphocytes, which tend to decrease in COVID-19, with the degree of decrease predicting disease severity.96–98 Since neutrophils, conversely, increase with disease severity, the neutrophil-to-lymphocyte ratio may be predictive as well.68 96 Reactive lymphocytes can also be identified by hematology analyzers, and were found to be lower in COVID-19 relative to other viral infections.99 Finally, NK cell, monocyte, and DC frequencies can be reduced in acute COVID-19.100
Within the lymphoid compartment, suppression of total CD3+ T cells, and particularly CD8+ T cells, has been shown to predict severe disease.101 Furthermore, a highly differentiated and/or exhausted CD8 +T cell compartment, which can result from long-term CMV infection, may predispose to worse COVID-19 outcome.40 102 103 Mazzoni et al104 found that polyclonal T cell activation revealed decreased cytokine production along with terminal differentiation in patients with COVID-19 generally. They also found reduced NK cell granzyme B expression in ICU patients specifically. Interestingly, the latter was correlated to serum IL-6 levels and was reversed by tocilizumab (anti-IL-6) therapy.
B cell subsets and plasmablasts may also be correlated with severe COVID-19.105 106 Specifically, there is an inverse relationship between the frequency of memory B cell subsets and symptom duration in recovered individuals.106 However, Bernardes et al105 found proliferating, metabolically active plasmablasts to be increased in severe disease, along with circulating megakaryocytes and increased erythropoiesis.
Many other studies have observed changes in frequency or phenotype of other specific immune cell populations, too numerous to review in detail here. As an example, Kulkarni-Munje et al107 noted a progressive decrease in pDCs and an increase of IFNγ-producing NK cells in severe disease relative to mild disease and healthy controls, while CD16 +monocytes trended higher in both mild and severe cases relative to controls. However, this latter finding may be due to increases in intermediate (CD14+CD16+) monocytes, as the frequency of non-classical (CD14lowCD16+) monocytes has been found to decrease with severe disease.57 Kulkarni-Munje et al107 also found an increase in activated (HLA-DR+, CD38+) CD8 +T cells along with IL-2-procucing CD4 +T cells and CD4+ T follicular helper cells in severe relative to mild disease. These latter could be identified by both CXCR5 and PD-1 expression.
The complexity and dynamic nature of the immune response makes it difficult to find a single cellular parameter that stably and unequivocally predicts disease course. In fact, patients are heterogeneous, and the time of sampling is important for many markers. Some, like lymphocyte and CD8+ T cell count, are relatively early and relatively consistent predictors, while others may be useful only in subsets of patients. Mathew et al108 identified ‘immunotypes’ of patients with COVID-19 based on different key features (like T cell activation and plasmablast levels) and their immune trajectories. Three of these immunotypes were linked to poor outcomes. Similarly, Lucas et al109 identified early immune signatures, including elevated cytokines and a progressive type 2 response (IL-5, IL-13, eosinophils, and IgE) that corresponded to a severe disease course. Rodriguez et al110 describe similar longitudinal changes including basophil and eosinophil increases in a longitudinal analysis of 39 severe patients with COVID-19. Single-cell sequencing to define longitudinal immune trajectories has also been done to compare COVID-19 with influenza.111 This study found distinct signaling pathways, including STAT1 and IRF3, active in COVID-19, with STAT3 and NFκB signaling active in influenza. Interestingly, both diseases showed increased IL-6 concentration, but patients with COVID-19 had a relative increase of IL-6 receptor gene expression in T cells and DCs. Finally, one of the most comprehensive longitudinal, multi-omic studies of COVID-19 was performed by Su et al43 who integrated plasma proteomics and metabolomics, PBMC transcriptomics, PBMC phenotyping, immune receptor repertoire sequencing, single-cell secretome, and electronic health record data. They describe an immunological shift between mild and moderate/severe cases, characterized by increased inflammation, decreased blood nutrients, and changes in immune cell populations. These latter include some of those described in other studies above, such as HLA-DRlow monocytes, exhausted CD4+ and CD8+T cells, and cytotoxic CD4+ T cells.
From all the above, it is obvious that there are numerous changes in COVID-19 in both antigen-specific and global populations of many immune cell subsets. Some of the simplest and broadest changes are actually reasonably good at predicting disease severity, including total lymphocyte and CD8 +T cell counts (both lower in severe cases), or neutrophil-to-lymphocyte ratio (higher in severe cases). These measurements can be made from a complete blood cell (CBC) test or (for CD8+ T cells) by clinical flow cytometry panels (BTNK assays). As such, these can already be implemented into clinical decision making.
More sophisticated measurements require more detailed multiparameter flow cytometry or CyTOF panels, including activation and differentiation markers for multiple cell types. These panels may also include activation with specific SARS-CoV-2 antigens (often overlapping peptide pools) to read out antigen-specific T cells; or global activation, for example, with phorbol myristate acetate +ionomycin, to read out global immune competence.112 Even higher information content can be achieved with single-cell transcriptomics, frequently in combination with TCR or Ig repertoire targeting and/or protein targeting with nucleic acid-tagged antibodies (AbSeq or CITE-seq). But these technologies are most practical for initial pilot investigations. Larger follow-up studies can then employ targeted flow or CyTOF panels with the most interesting markers gleaned from single-cell transcriptomics.
Other clinical monitoring tests
Other clinical markers that are potentially useful along with monitoring of immune cells, antibodies, and cytokines, include markers of inflammation and tissue injury, such as C reactive protein (CRP), procalcitonin, or cardiac troponin. Tan et al evaluated multiple such factors for their ability to stratify patients for severity or mortality.113 In addition to IL-6, lymphocyte percentage, and viral load, this study found CRP and procalcitonin to be most sensitive and reliable in separating survivors from non-survivors. Lymphocyte percentage was most sensitive at discriminating the full spectrum of severity and outcomes. Majure et al further showed that hospitalized patients with COVID-19 with elevated troponin have a greatly increased risk of mortality.114 For information on even more clinical tests in this disease, a review of 72 studies of clinical laboratory markers for their ability to predict COVID-19 disease severity and outcome is given here.115 Interestingly, some systemic inflammatory markers remain elevated 40–60 days postinfection, even in mild or asymptomatic patients.116
Autoantibodies have recently been associated with COVID-19, including antibodies to phospholipids and phospholipid-binding proteins,117 118 antibodies to dsDNA and other specificities associated with systemic autoimmune diseases,119 and the already-mentioned antibodies to IFNα.70 Furthermore, Woodruff et al120 found that severe COVID-19 was associated with extrafollicular B cell development, oligoclonality, and autoimmune clonotypes that were also seen in systemic lupus erythematosus patients. Together, these studies suggest autoimmune features to be a frequent occurrence in severe cases of COVID-19.
SARS-CoV-2 viral load is another clinical parameter that can be monitored. Viral load can be quantitated in plasma, which is positive in a minority of hospitalized patients and rarely in non-hospitalized patients. Fajnzylber et al98 found that plasma viremia was associated with disease severity and with mortality. Zheng et al121 also identified SARS-CoV-2 RNA in stool, as well as serum and respiratory samples. They showed a longer duration of virus positivity in all sample types with severe disease.
Sex differences
Males tend to have worse COVID-19 outcomes than females, and this may be due in part to immunological differences. For example, males tend to have more robust production of innate cytokines and non-classical monocytes; while females have more robust T cell responses.122 Interestingly, poor T cell responses were associated with worse outcomes in male patients, but not females; while innate cytokines were correlated with worse outcomes in females but not males.122 Thus, some of the immune correlates of disease severity may in fact be sex-specific, meaning that clinical decisions based on these tests need to factor in the patient’s sex.
Biomarkers of long COVID-19, MIS-C, or organ damage
The ability to predict and hopefully intervene to prevent development of ‘long COVID-19’ in adults or MIS-C in children would be highly useful. Elevated inflammatory markers and cytotoxicity genes, as well as autoantibodies, have been seen in MIS-C123 and some of these may be found to be predictive factors. The U.S. Tri-State Pediatric COVID-19 Research Consortium found that non-Hispanic Black children were over-represented among MIS-C relative to respiratory disease cases. This consortium further found that low absolute lymphocyte count and high CRP were predictors of severe MIS-C. I am as yet unaware of similar studies to find immune predictors of patients who will develop ‘long COVID-19’ or postacute COVID-19 syndromes. Similarly, prediction of specific organ damage in patients with COVID-19 has so far been limited to acute monitoring of markers of tissue injury (eg, cardiac troponin I and N-terminal pro-brain natriuretic peptide for cardiac damage). And coagulopathies are monitored only by coagulation-specific markers such as D-dimer.124
Current and future clinical monitoring studies
Large longitudinal studies such as IMPACC (https://www.niaid.nih.gov/clinical-trials/immunophenotyping-assessment-covid-19-cohort-impacc) and similar cohort studies in Europe, Australia, and Africa will soon provide a wealth of viral and immune monitoring data on patients treated in those regions of the world. For children, COVID-19: Pediatric Research Immune Network on SARS-CoV-2 and MIS-C study https://clinicaltrials.gov/ct2/show/NCT04588363) will immunophenotype MIS-C samples to better understand this rare complication of pediatric COVID-19. With standardized methods, these studies could help to uncover geographical and other factors related to disease severity and sequelae, as well as provide corroborative data across different populations. Also of interest would be studies that could identify biomarkers of susceptibility and severity of COVID-19 in healthy individuals prior to infection, a more difficult area of study that is as of yet largely unexplored.
Conclusions
The literature on immunity to SARS-CoV-2 is already so vast that it cannot be comprehensively cited in a single review. Still, the studies highlighted here point to aberrations of both innate and acquired immunity in the setting of severe or fatal COVID-19. In blood, these include elaboration of high levels of IL-6 and other proinflammatory cytokines; proliferation of neutrophils (and developing neutrophils); reduction of lymphocytes, especially CD8+ T cells; strong elevation of plasmablasts; development of HLA-DRlow monocytes and decrease of non-classical monocytes; appearance of MDSCs; decreases in frequency and function of pDCs; and numerous changes in other immune cell subsets and their phenotypes (table 1). While virus-specific antibodies and T cells are produced in the vast majority of cases, the clonality and epitope specificity of these adaptive responses may provide clues to the disease course. In addition, clinical tests for soluble factors such as CRP, procalcitonin, and troponin can help inform the trajectory of disease. These, along with CBC and/or TBNK flow cytometry are readily accessible to clinicians currently, and they should be used to identify patients needing the most careful monitoring and early intervention. More sophisticated tests may eventually also find clinical utility, especially if they can discriminate patients who develop particular symptoms and sequelae, or if they identify individuals at risk of worse outcomes even prior to infection or vaccination.
COVID-19 biomarkers by platform and regulatory approval level
References
Footnotes
Contributors HTM wrote and edited the manuscript.
Funding The authors have not declared a specific grant for this research from any funding agency in the public, commercial or not-for-profit sectors.
Competing interests No, there are no competing interests.
Provenance and peer review Not commissioned; externally peer reviewed.